Une protéine dans la soupe ? (2e partie)

Résumé : Sous mode d’une conversation, nous poursuivons ici la description des étapes d’un processus biologique éminemment banal et qui se produit constamment au sein de nos cellules : la fabrication d’une protéine. Une « simple » cellule fonctionne déjà comme un être vivant complet, capable de synthétiser des molécules à partir d’énergie et de molécules préexistantes. Encore faut-il un site adéquat, l’information requise et des outils de contrôle, les messagers. Or cette complexe machinerie, nécessitant un plan de montage et le transfert de matériaux, se réalise en un temps très court selon une suite ordonnée d’opérations (épissage, traduction). La machinerie de traduction peut polymériser plus de 10 pièces par seconde ! À aucun stade ne peut intervenir un quelconque « hasard ».
Dimanche après-midi. Le temps rêvé pour de bons moments en famille. C'est cet instant que choisit Jean pour reprendre sa conversation avec son papa Louis.
J : Dis, papa ! je comprends bien que, sans un agent extérieur, je peux attendre en vain que le sac plastique devienne un Playmobil, mais le temps ne pourrait-il pas jouer sur la mobilité moléculaire du peptide pour que ce dernier donne finalement une protéine ? Tu m'as indiqué que le reploiement des molécules se faisait, presque automatiquement, en fonction de la propriété des atomes. Pourquoi y aurait-il plus de différences entre un petit peptide apparu « spontanément » et une protéine, qu'entre un sac plastique multicolore et un personnage de Playmobil ?1
L : Il y a plus de différences au sens où la protéine, du fait d'une capacité à être mue par son milieu ambiant, peut se présenter sous différentes formes. En biochimie, des expériences sont réalisées pour déterminer le pH et la température donnant l'activité optimale de protéines appelées enzymes. Cela est une illustration de ce que je viens de te dire : une variabilité de conformations définies d'avance.
L'analogie et la comparaison entre le Playmobil et la protéine, qui comportent tous deux une certaine complexité tridimensionnelle, voulaient illustrer la nécessité d'une cause efficiente, ou agent – comme tu l'as très bien dit –, extérieurs pour passer éventuellement d'un terme à l'autre. Éventuellement, car il faut être en puissance de le devenir.
Ni le temps ni le hasard (événement arrivant par définition rarement) ne peuvent causer par eux-mêmes ce passage bien que « les briques élémentaires » soient semblables : du plastique dans un cas, des acides aminés dans l'autre cas. Je vois mal comment un sac plastique pourrait se transformer en Playmobil sinon par une intervention humaine qui fond et réutilise le premier pour tenter d'obtenir le second. De même, un peptide et une protéine active ne se différencient pas que par leur longueur de chaîne et la complexité finale de structure. Pour obtenir une protéine, de nombreux agents internes de la cellule interviennent dont des protéines !
J : La cellule ?
L'élaboration d'une protéine se fait dans le cadre d'un organisme vivant et d'une finalité. Elle est comme un outil qui participe à la vie d'une entité : la plus petite est la cellule. La bactérie ou microbe, un être unicellulaire qui a pourtant déjà la complexité d'une usine, en est un exemple.
La vie, c'est un constat d'expérience. Un animal et son cadavre sont très semblables : organes, cellules, atomes sont présents. Si l'animal cesse d'être animé, il cesse d'être vivant. Vivre, c’est agir de façon immanente. L’immanence implique deux parties dans l’action : l’action doit procéder d’un principe intrinsèque agissant ; et l'action doit se terminer dans l’agent lui-même.
J : Un être unicellulaire ?
L : Un organisme composé d'une seule cellule, petite usine qui a tout pour fonctionner par elle-même si elle est approvisionnée par un milieu ambiant favorable à sa survie. Les conditions à réunir pour avoir une cellule vivante sont nombreuses, car la vie cellulaire se caractérise en particulier par les synthèses de molécules.
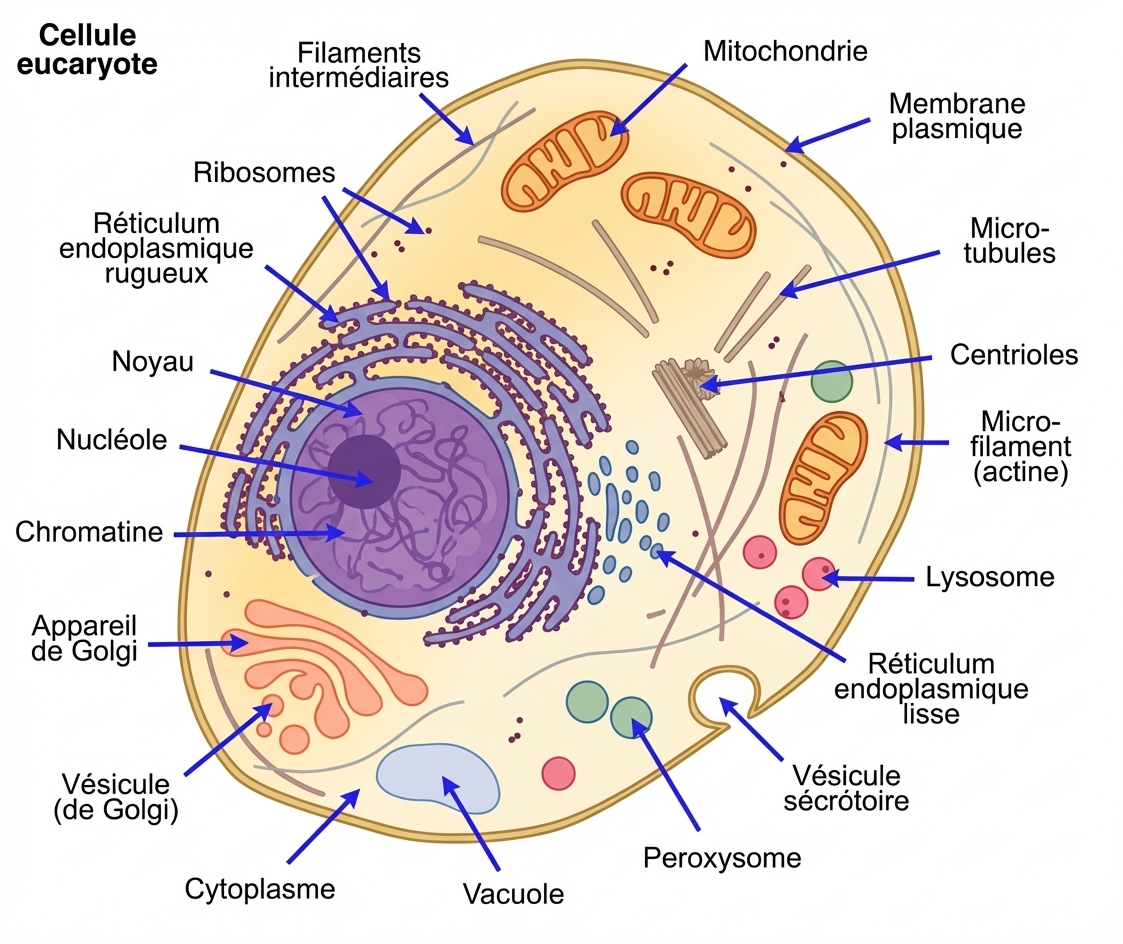
Schéma d'une cellule.
J : Conditions nombreuses ? Cela me fait penser à la notion de complexité irréductible2 de Michael Behe ?
L : Oui, et à une Intelligence conceptrice.
Toute synthèse cellulaire :
nécessite une matière première, sous la forme de molécules simples ou précurseurs – tes briques élémentaires –, et d’énergie utilisable (nucléosides triphosphates appelés ATP et GTP ;
se réalise dans un site précis où se trouvent les outils d’assemblage (enzymes, etc.) ;
requiert une information, directement via des molécules informationnelles pour les macromolécules séquencées (acides nucléiques et protéines), et indirectement par le biais des enzymes ;
est contrôlée par des messagers, des protéines qui, soit se fixent sur des récepteurs du plasmalemme (ou membrane plasmique) et sont relayées par d’autres messagers intracellulaires (seconds messagers), soit exercent directement leur action sur le génome (messagers stéroïdiens et hormones thyroïdiennes).
L'homme utilise les connaissances acquises pour manipuler, transformer le vivant mais sans jamais le produire. Les protéines artificielles sont réalisées par mimétisme des processus observés dans le vivant et par transformation chimique.
J : Tu as dit que la synthèse d’une protéine nécessite des protéines. En faut-il beaucoup ?
L : Oui, de nombreuses, sans compter l'ADN.
In vivo, les protéines fabriquées ne sont pas le résultat de la jonction aléatoire d’acides aminés baignant dans une soupe cellulaire. Toute une machinerie cellulaire entre en jeu à chaque fois.
La première étape dans la synthèse d’une protéine est l’élaboration ou plutôt la recherche du plan de construction. Ce plan est codé dans une molécule appelée ADN. Un gène est comme un plan d'assemblage de la structure primaire d'une protéine. Chaque cellule possède un catalogue complet des gènes nécessaires à l’espèce. Chez les êtres pluricellulaires, pour telle cellule prise individuellement, seuls certains sont accessibles. Ceux qui sont dans les zones de condensation de la chromatine3 ne le sont pas.
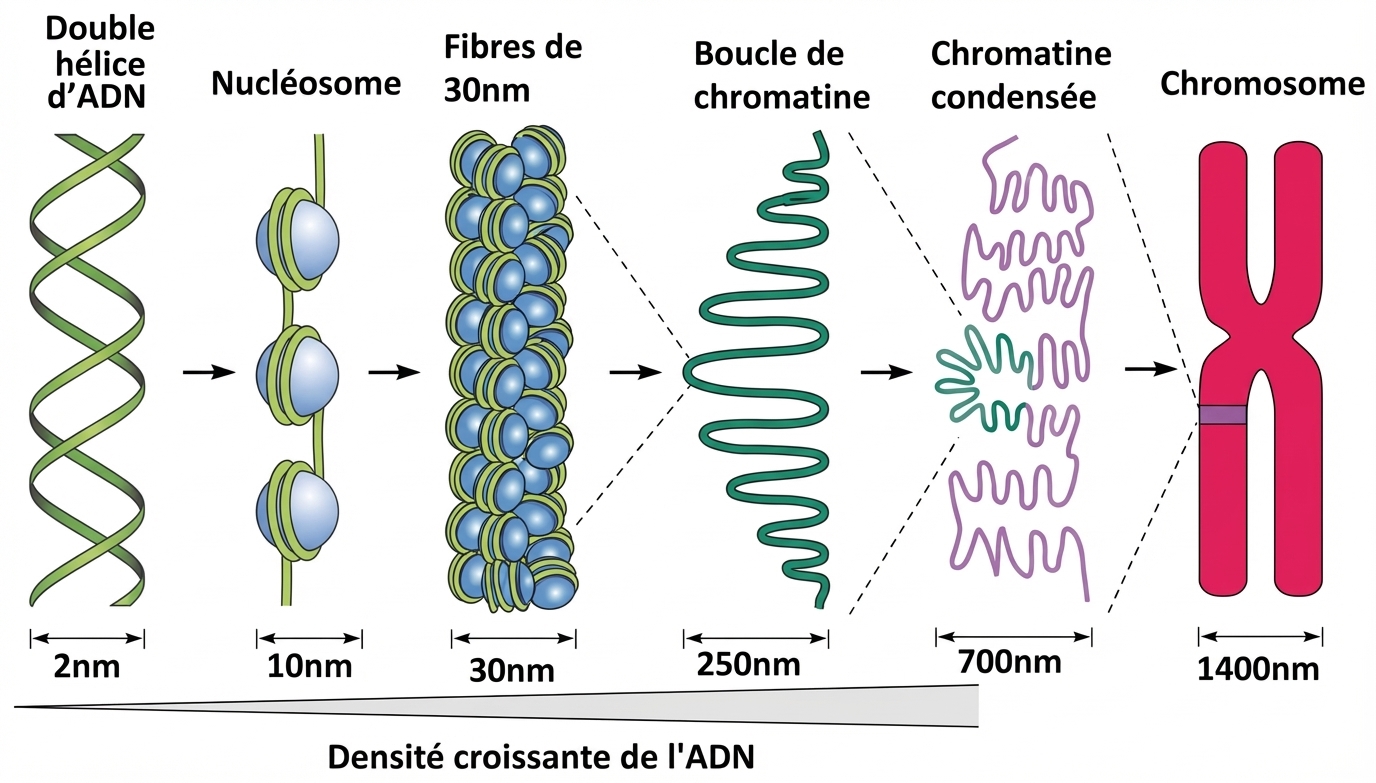
Chromatine et ADN : comment densifier au mieux une immense quantité d’information.
J : Pourquoi la chromatine est-elle condensée par endroits ?
L : À cause de toute l'histoire de l'organisme. Depuis la première cellule œuf, de nombreux événements se sont produits dont ont résulté des expressions inégales de gènes et des différenciations cellulaires. La régionalisation a permis une répartition des tâches et une expression différente du contenu du génome : chaque cellule n'utilise pas tous les gènes ou plans.
J : Il y a donc des différences entre une cellule de microbe et une cellule d'homme ?
L : Oui, plusieurs ; mais nous y reviendrons peut-être une autre fois.
Pour obtenir une protéine donnée, l’accessibilité « chromosomique » du plan de montage est donc le premier défi à relever. Il est résolu par l’histoire, programmée, de la cellule.
Le second défi est la mise en relation de ce plan avec la chaîne de fabrication.
Problème majeur chez les êtres pluricellulaires : l’enveloppe nucléaire est une barrière infranchissable pour le volumineux ADN qui contient l’information codée dans le noyau cellulaire. Il ne peut passer dans les pores nucléaires de 40 microns. L’ingénieur divin a donc prévu la réalisation d’un courrier, une copie de l’instruction de montage : l’ARN messager dit ARNm.
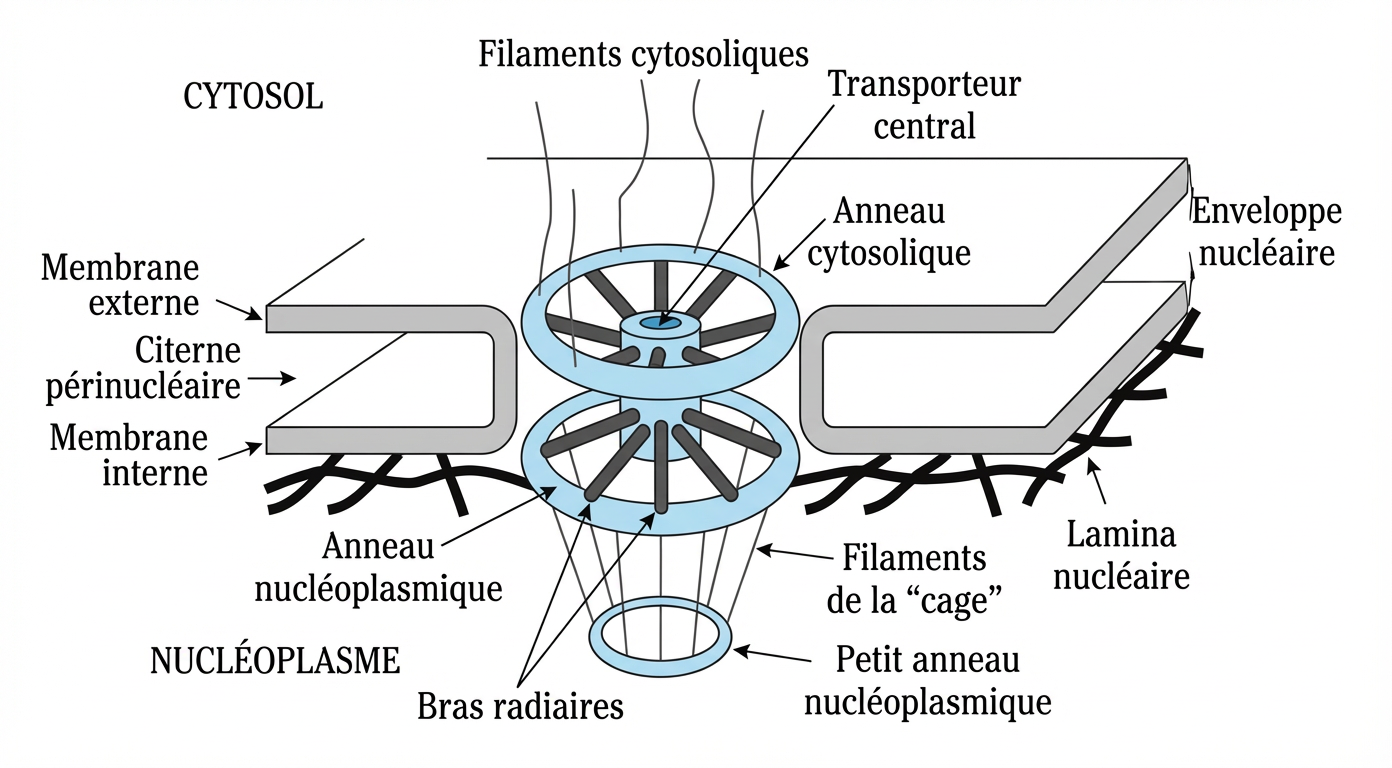
Pore nucléaire dans la membrane.
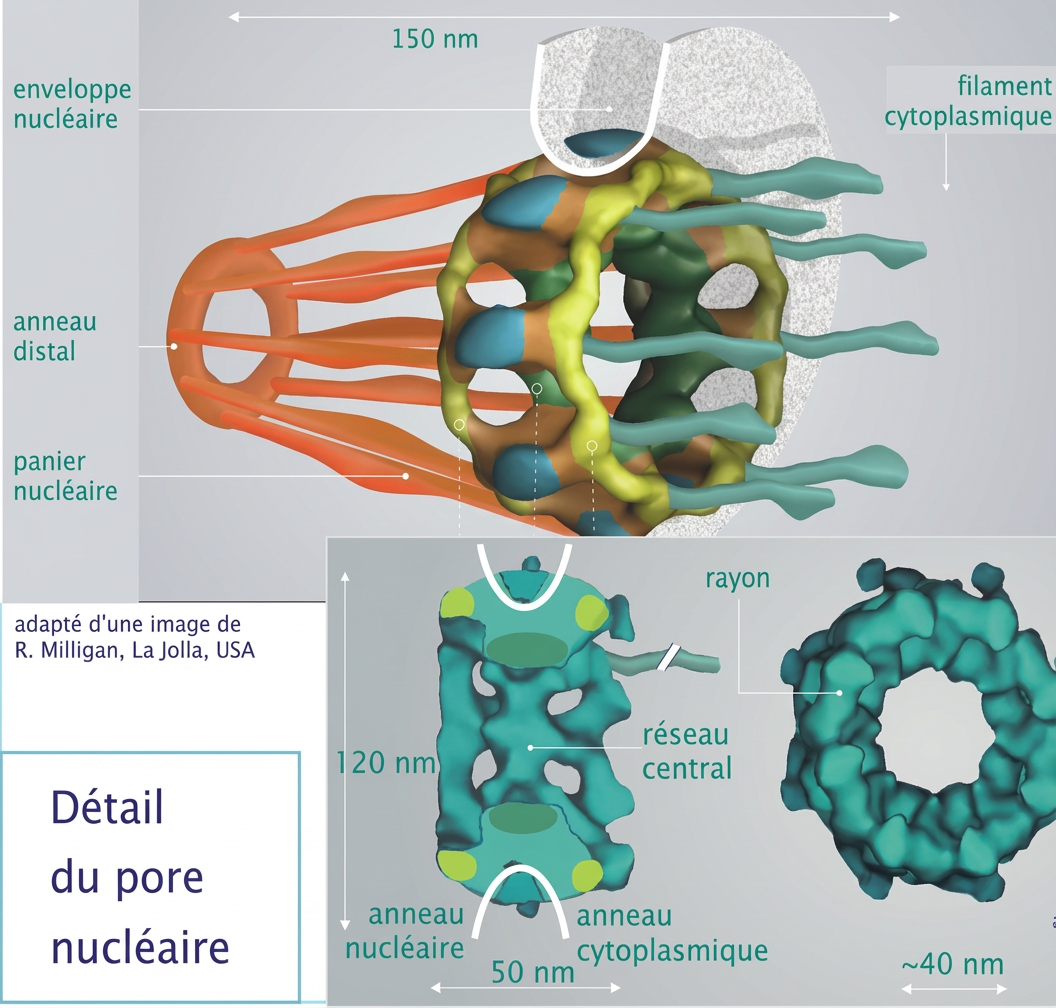
Détail du pore nucléaire.
La réalisation de la copie du message, appelée transcription, en vue de la fabrication d’une chaîne d’acides aminés, par un processus appelé traduction, requiert une combinaison de différentes protéines : l’enzyme ARN polymérase II, des facteurs de transcription généraux et spéciaux ainsi que leurs facteurs associés.
Il est difficile d’évaluer numériquement le nombre d’intervenants à la réalisation de cette copie qui est appelée à ce stade : ARN pré-messager. Cette copie subit, en effet, des modifications qui ont deux objectifs principaux :
favoriser le transfert de l’ARN par les pores (trous) du noyau et protéger l’information : l’addition d’une coiffe et d’une queue poly-A ont ce rôle.
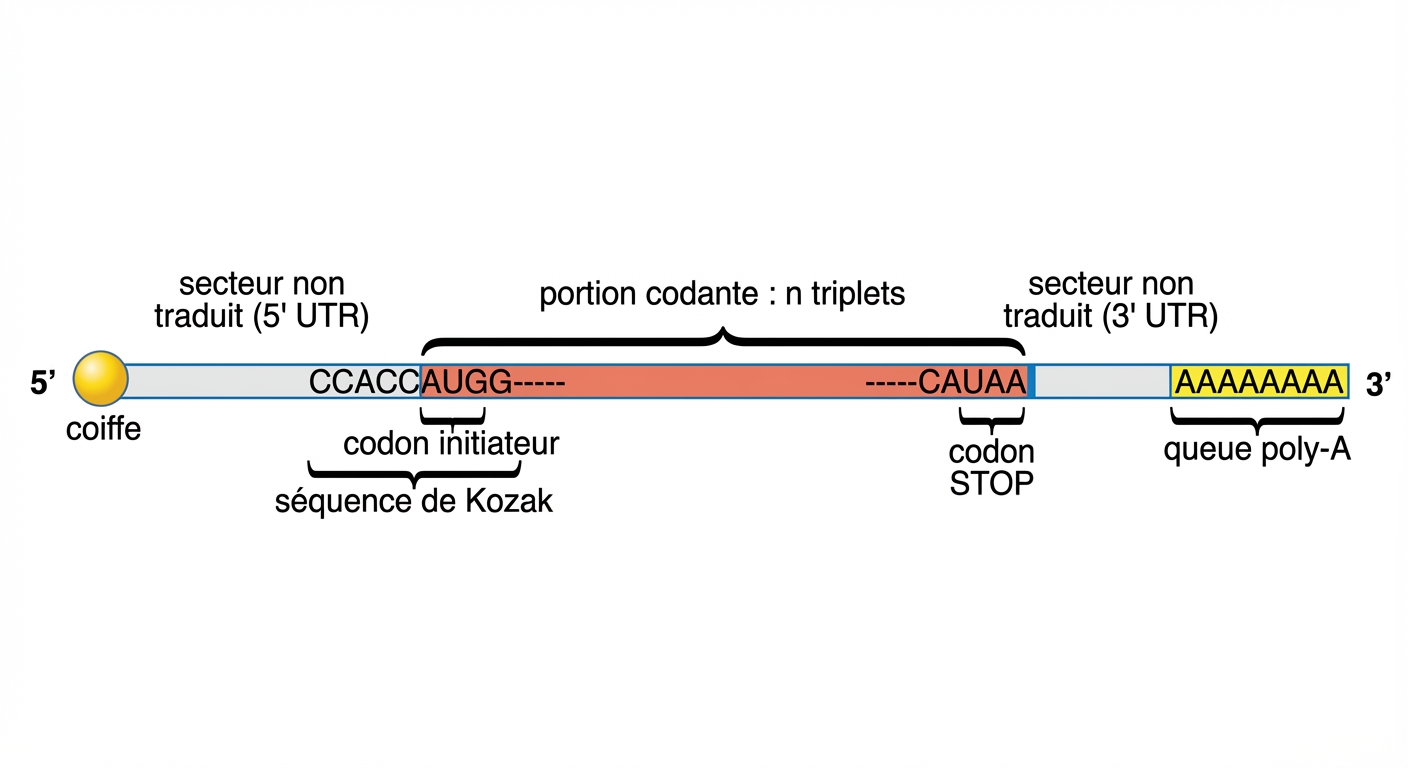
Organisation d'un ARNm eucaryote4.
Bien qu’il soit protégé, la demi-vie de l’ARNm oscille entre quelques minutes et quelques heures. C’est heureux. La régulation fine de la production le demande. Les causes de cette brièveté sont diverses : activité dégradatrice d’enzymes, les exonucléases ; hydrolyse par un complexe enzymatique ; protéines régulatrices qui se fixent sur les parties non codantes de l’ARNm ; micro-ARN interférents, etc.
J : Demi-vie ?
L : On parle de demi-vie pour indiquer le temps au bout duquel la moitié des copies ARNm sont détruites.
Nous ne pouvons qu’admirer ce fin équilibre qui permet d’obtenir une production qui ne soit pas une surproduction encombrante pour la cellule et nuisible pour l’organisme.
Il suppose aussi communication entre les cellules…
Une dernière modification chez les êtres pluricellulaires, l’épissage [mot tiré du néerl. splitsen : « attacher deux cordages par entrelacement »] des exons, permet souvent aux génomes de produire des protéines différentes à partir des mêmes gènes.
J : Pourrais-tu expliquer un peu ce qu'est l'épissage ?
L : L'épissage est une trouvaille du Créateur pour stocker beaucoup d'informations sur une petite surface de bobine.
Chez les eucaryotes (organismes à noyau), l’épissage est un processus par lequel les ARN transcrits à partir des gènes de l'ADN peuvent subir des étapes de coupure et ligature contrôlées qui conduisent à l'élimination de certaines régions dans l’ARNm final. Les segments conservés s’appellent des exons et ceux qui sont éliminés s’appellent des introns. Souvent les gènes codant des protéines, ainsi que certains gènes d’ARN non codants [ par exemple ceux des ARNt ou de transfert, ARN indispensables pour la fabrication ], sont constitués d’une suite d’exons et d’introns alternés. Cela permet, à partir d’une même séquence d’ADN, d’obtenir différentes protéines par la simple sélection de morceaux choisis (les exons). Il est probable que la composition du splicéosome favorise telle ou telle combinaison.
Lors de la transcription ou copie du gène codant par exemple l’insuline, un ARN pré-messager est synthétisé. Il est épissé dans le noyau de la cellule pour donner l’ARN messager dit mature.
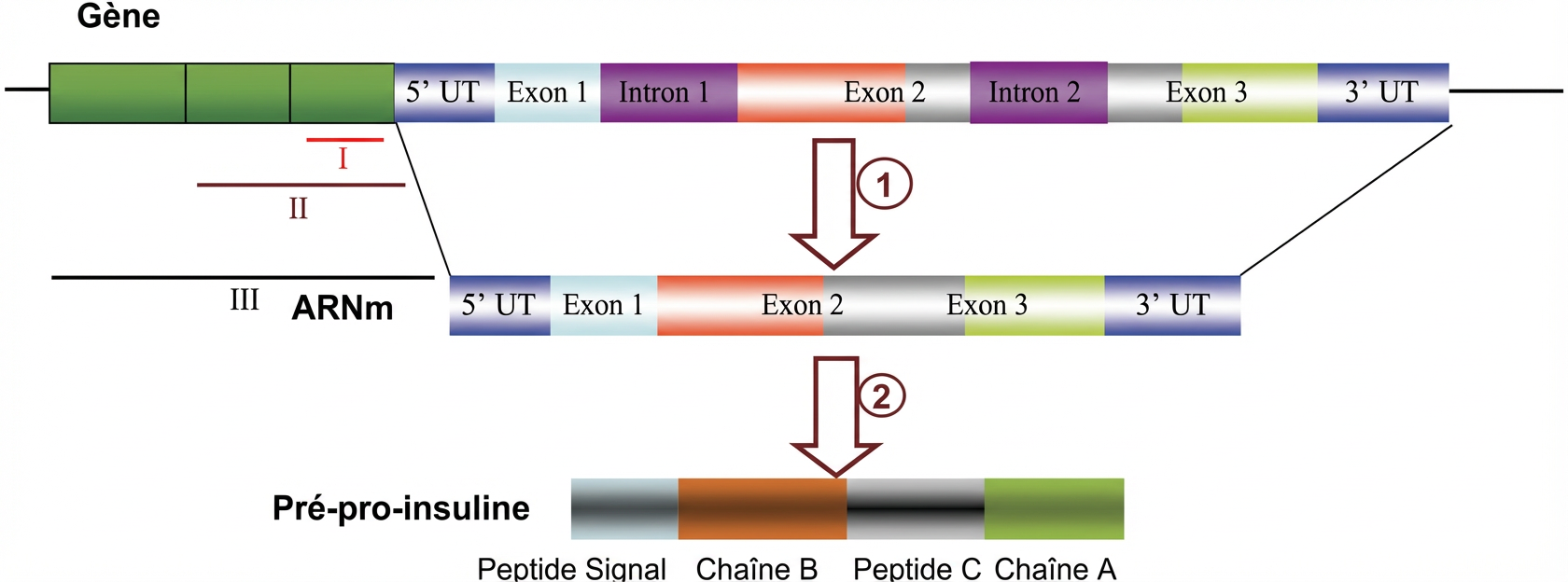
Gène INS, son ARNm après épissage et le polypeptide résultant de la traduction.
1/ transcription et épissage du gène codant l’insuline ; 2/ traduction en une chaîne d’acides aminés.
L'épissage est beaucoup plus long que la transcription, cette dernière prenant quelques minutes contre environ une heure et demie pour l'épissage.
Splicéosome.
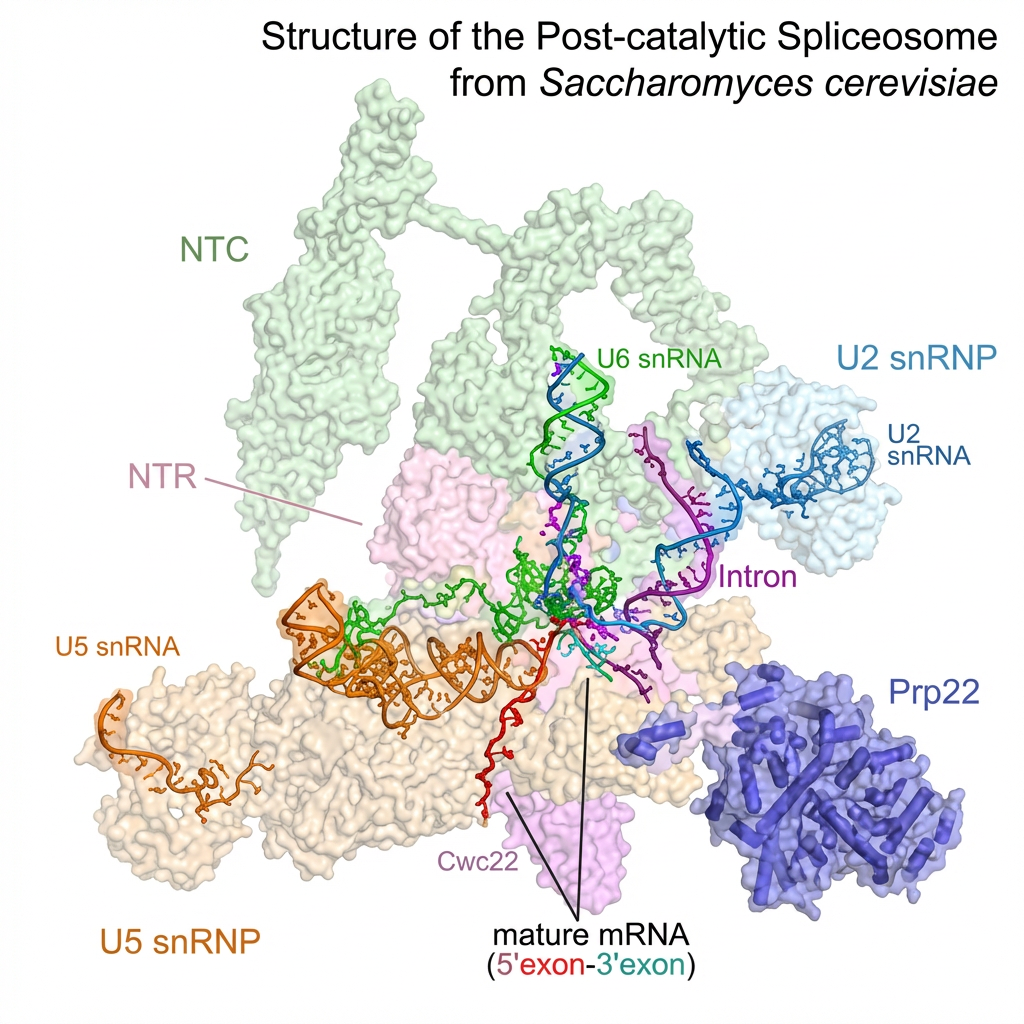
Schéma d'un splicéosome de levure de bière.
J : La composition du… splicéosome ?
L : Un splicéosome (épissage se disant splicing en anglais) est le nom collectif donné à un ensemble de complexes ribonucléoprotéiques d’environ 25 nm de diamètre, qui catalysent l’épissage. C'est en quelque sorte « la machine à épisser ». Ils sont constitués de petits ARN nucléaires de 100 à 300 nucléotides riches en uracile associés à des protéines (les snARN ou small nuclear ARN, ARN non codants). Les peptides qui en résultent ayant des propriétés très particulières, on voit mal comment les combinaisons ne seraient pas pensées d’avance.
J : ARN non codants ?
L : Il s'agit de tous les ARN qui ne sont pas destinés à être décodés lors de la traduction pour donner une séquence d'acides aminés, séquence préliminaire à l'élaboration d'un polypeptide ou d'une protéine.
Alexandra Henrion Caude, une spécialiste des ARN, en fait une brève énumération dans son livre Les Apprentis sorciers : microARN (20 lettres, régule un gène) ; ARNr et ARNt, qui participent à la traduction ; ARNsi ; ARNsh ; ARNpiwi ; ARNe ; ARNnc ; ARNsn ; ARNsno ; ARNsca ; ARN hôtes d’autres ARN ; ribozymes ; ARNcirc ; ARNvt ; ARNy. Les ARN provenant des introns en font partie. Il y a encore bien des merveilles à découvrir sur les ARN non codants.
J : Que se passe-t-il après l'épissage ?
L : Des mécanismes de contrôle s'assurent que les ARNm ont été correctement épissés avant de permettre leur exportation.
In fine, l’ARNm mature, constitué des seuls exons, est exporté vers le cytoplasme pour être traduit en protéine. Des transporteurs assurent son passage à travers les pores nucléaires.
J : C'est alors la traduction ?
L : Oui. À sa sortie du noyau, notre courrier ARNm arrive assez vite à entrer en contact avec les ribosomes situés à proximité.
La traduction consiste dans l’assemblage ordonné d’acides aminés dans le cytosol, le milieu liquide de la cellule.
Les ribosomes sont d‘énormes complexes d’environ 80 protéines et 4 molécules d’ARNr ou ARN ribosomiques associés à des ARN de transfert : les ARNt. Les ARNt, « adaptateurs » ou « outils de correspondance », sont chargés d’un acide aminé par 20 enzymes aminoacyl-ARNt-synthétases spécifiques.
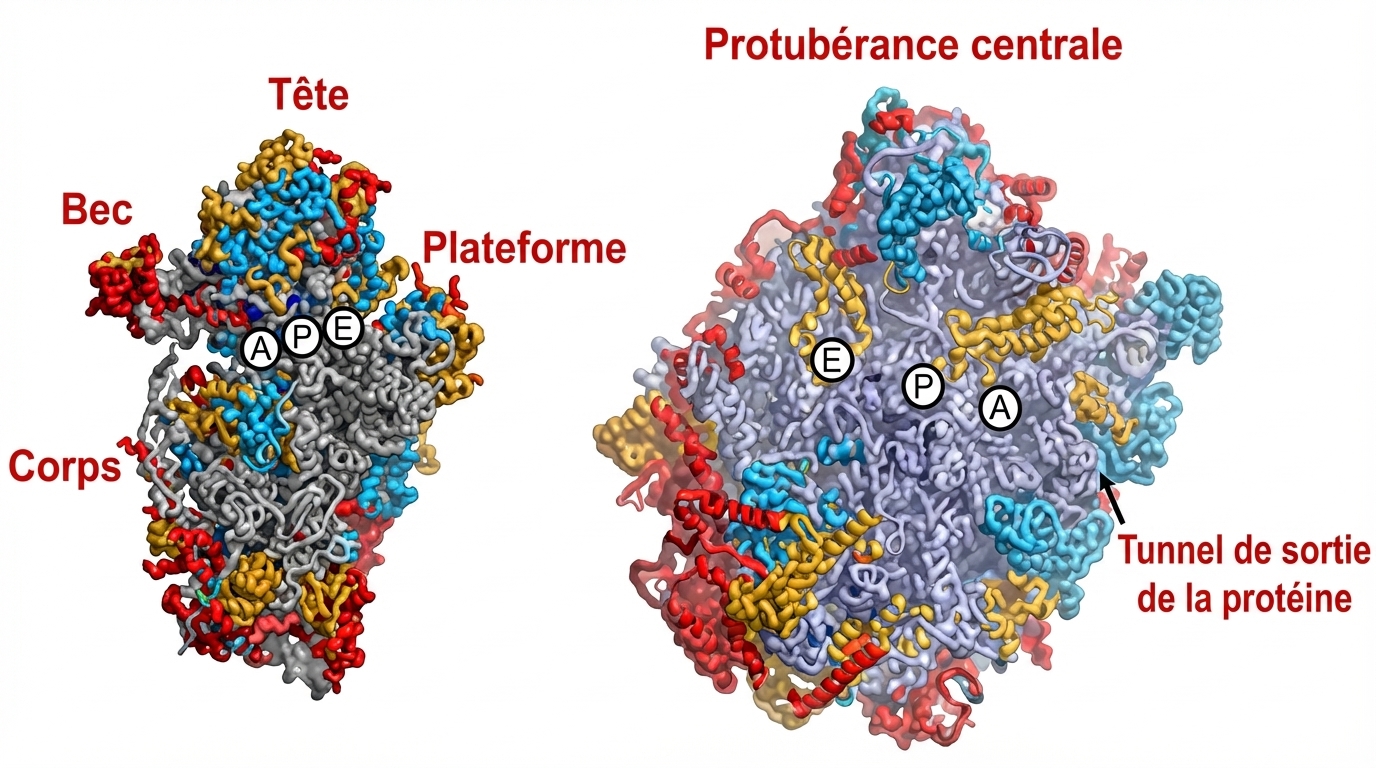
Ribosome vu de face et de profil.
Ces réactions de protéosynthèse sont consommatrices d’énergie et la présence intracellulaire de centrales énergétiques, les mitochondries, n’est pas accessoire.
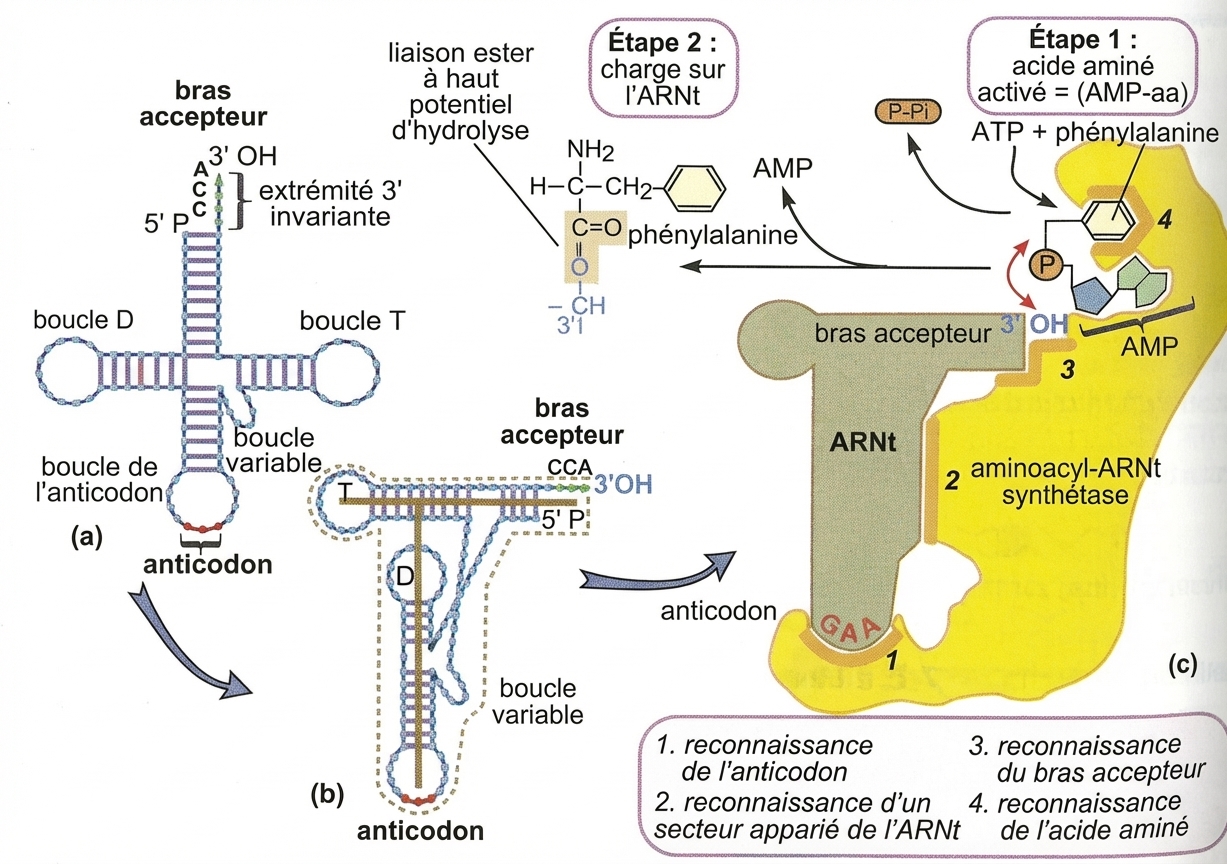
Structure secondaire (a) et tertiaire (b) d'un ARNt ; aminoacyl-ARNt-synthétase de la phénylalanine (c).
L'enzyme présente quatre sites de reconnaissance. Si le quatrième site est très spécifique à un acide aminé, le premier peut, lui, recevoir différents anticodons. C'est le site 2 qui met en relation une partie spécifique de l'ARNt à une des 20 enzymes alors que le site 3 est invariant. C'est au niveau de ce dernier que l'acide aminé est fixé à l'ARNt5.
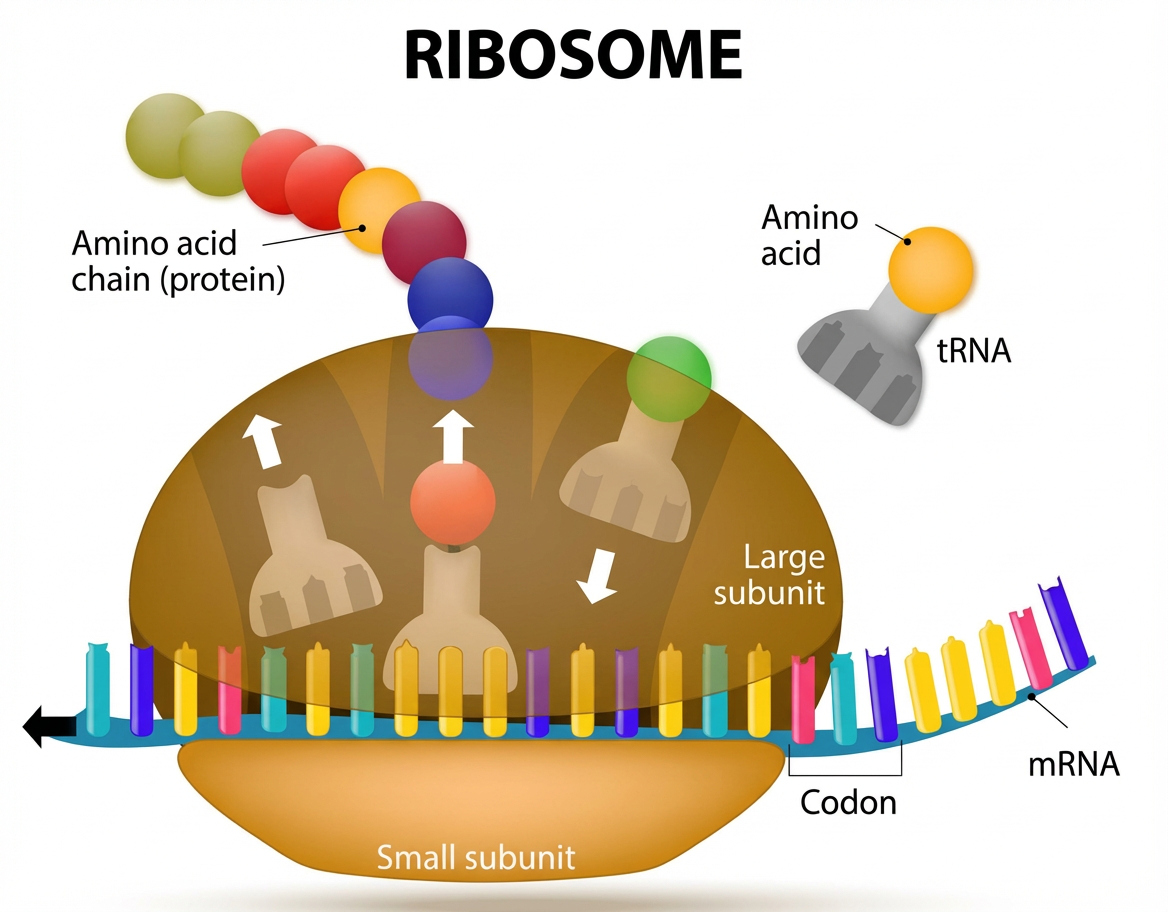
Traduction.
L'ARNt vient se fixer par son anticodon sur le codon de l'ARNm correspondant au sein du ribosome. Il ressort de ce dernier après avoir perdu l'acide aminé qu'il portait.
L’outil de correspondance, l’ARNt, est un adaptateur bipolaire. Il comporte à un pôle le triplet de nucléotides correspondant à un codon de l’ARNm qui, comme les roues dentées d’une montre à aiguille, va s’emboîter parfaitement6.
À l’autre pôle, l’ARNt porte l’acide aminé qu’a chargé sur un bras accepteur l’enzyme aminoacyl-ARNt-synthétase. Étant donné qu’il n’y a que vingt enzymes pour une quarantaine d’ARNt, c’est la même enzyme qui charge un acide aminé donné sur des ARNt isoaccepteurs, ce qui signifie une reconnaissance non absolue de l’anticodon dans ce cas. Divers contrôles fondamentaux sont exercés lors de cette étape. Certains acides aminés correspondront à plusieurs codons : c’est la redondance du code génétique. Ainsi l’ARNt s’adaptant moléculairement à UUU porte le même acide aminé que l’ARNt s’adaptant à UUC : la phénylalanine.
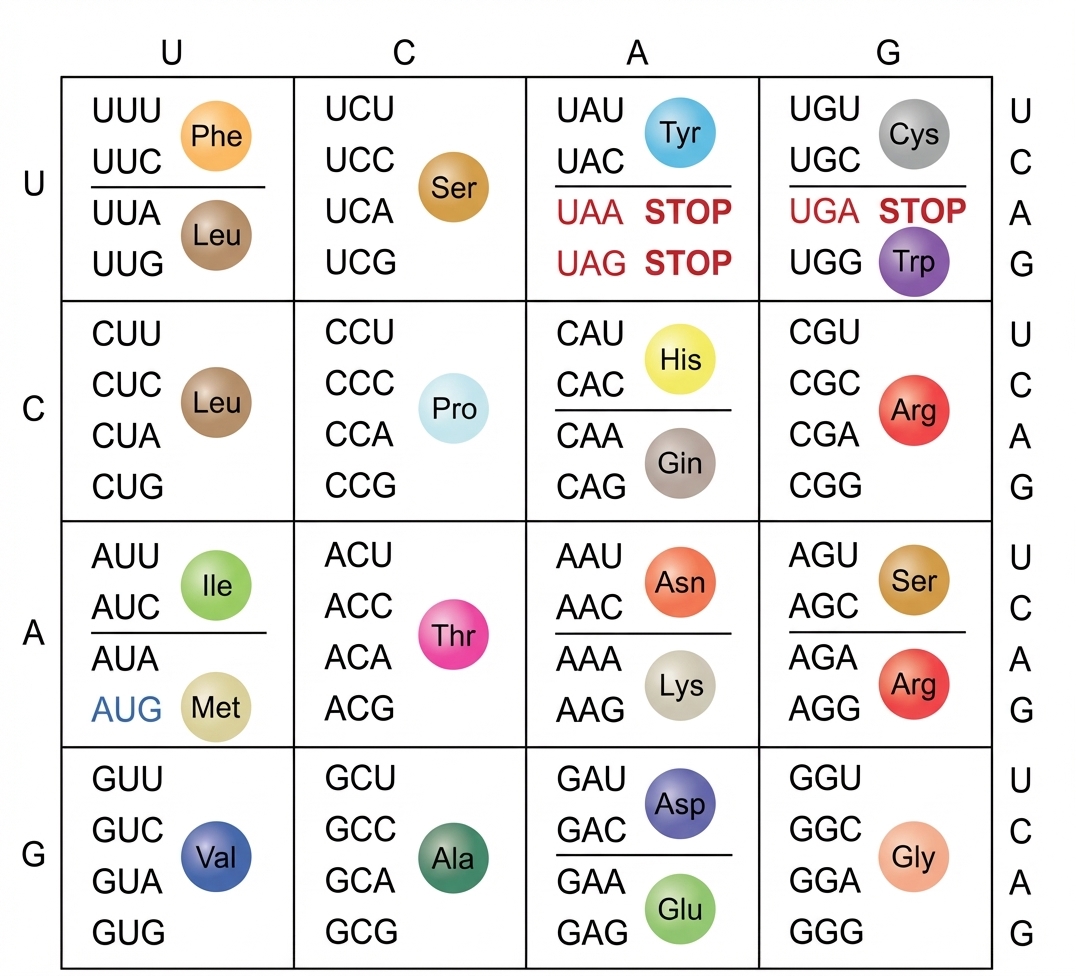
Correspondance codons-acides aminés.
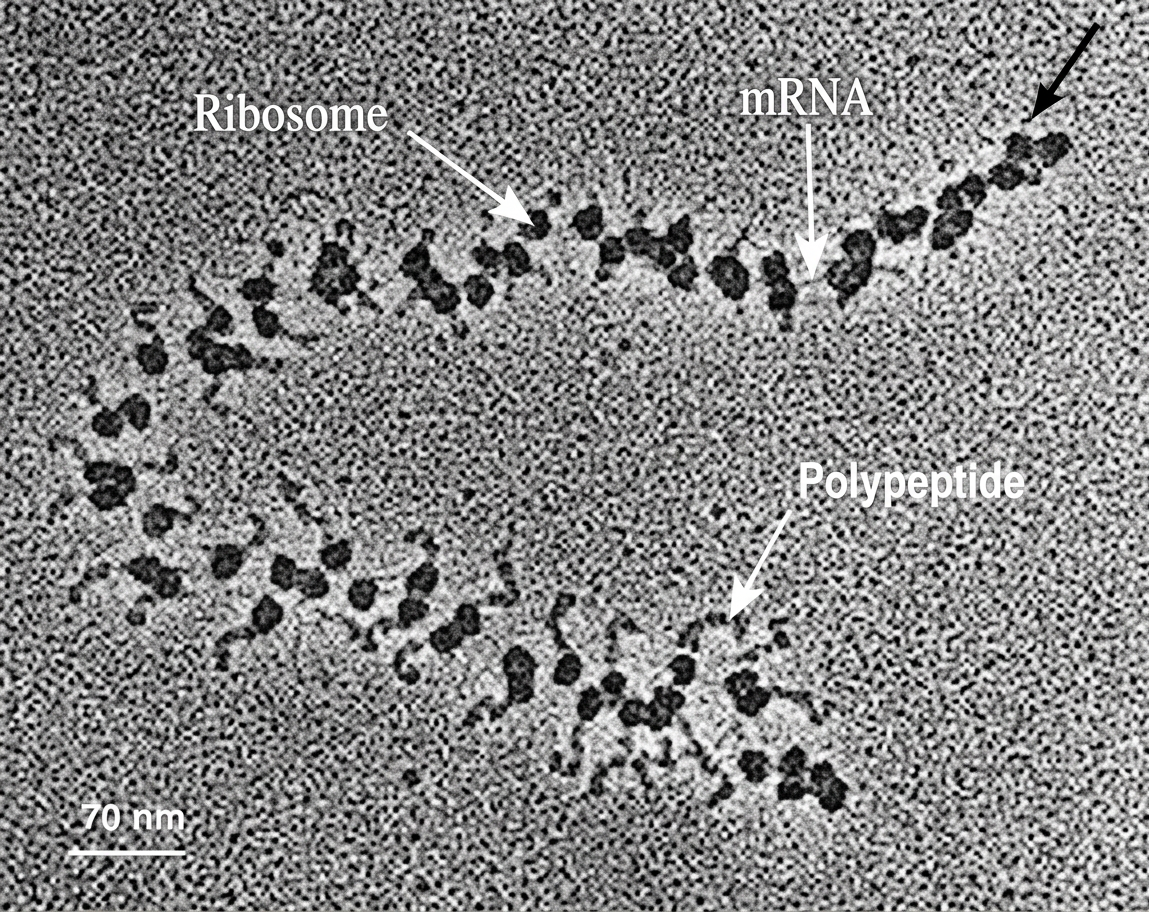
Traduction vue au microscope. Sur un seul ARNm plusieurs ribosomes (points noirs) se fixent successivement. La synthèse du polypeptide (chaînes latérales grises) s'achève à une extrémité de l'ARNm (bas de l'image) alors qu'elle débute pour le ribosome de l'autre extrémité (flèche noire).
Quelques codons n’ont aucun adaptateur et c’est heureux ! Ce sont des signaux de fin de traduction.
Par le nombre d’acteurs qu’elle met en jeu et la précision qu’elle requiert, la traduction est sans doute la plus complexe des synthèses macromoléculaires. La machinerie de traduction peut polymériser 10 à 20 pièces par seconde, processus dit « relativement lent » mais compensé par le fait qu’un même ARNm est traduit par de nombreux ribosomes qui travaillent indépendamment les uns des autres. La capacité de traduction dépend aussi de la quantité des divers facteurs associés disponibles [protéiques : IF ou facteurs d’initiation] et de la durée de vie de l’ARNm.
J : Le nombre d'acteurs intervenant est vraiment impressionnant. Ce sont des centaines de protéines qui sont nécessaires : splicéosome, ribosome, facteurs divers et variés, centrale énergétique faite de protéines…
L : Et ce n'est pas fini ! Nous n'en sommes qu'à la structure primaire. Il va falloir parler maintenant des modifications post-traductionnelles sans lesquelles le polypeptide ne sert à rien pour l'organisme.
Mais il est maintenant tard pour t'en parler. La prochaine fois nous éclaircirons tout cela en prenant l'exemple de l'insuline, qui fait défaut à ton oncle Henri, diabétique.
Chaque protéine a son rôle particulier et son utilité pour la vie de l'organisme. À ce jour, on ne sait pas précisément combien de protéines différentes existent. Chez l'homme, ce nombre avait été évalué à environ 20 000 il y a quelques années, puis 60 000. À présent, il n’est pas rare d'entendre des estimations dépasser les 100 000 protéines différentes. Cela te montre bien que la vie n'a pu apparaître par le jeu du hasard !
Bibliographie principale :
P. PEYCRU, D. GRANDPERRRIN & C. PERRIER, Biologie tout-en-un - BCPST 1re année, Malakoff, Dunod, 2017, ch. 1-10, p. 17-19.
Cet article est publié par le Centre d'Études et de Prospective (CEP)
Voir l'article sur le site